|
Bottom-up regulation of multitrophic foodwebs
While working in the lab of Anurag Agrawal at Cornell University, my work has focused on the common milkweed (Asclepias syriaca) as a model system. Common milkweed has large genetic variation in two potent defensive traits, sticky latex and a group of toxic chemical compounds known as cardenolides. Nonetheless, milkweed has a large community of specialized herbivores, including four species of aphids with varying degrees of host specialization and different life history traits. I have used this model system to study the selective forces that structure natural populations, and especially how genetic variation in plants can directly affect herbivores, or indirectly affect the interaction among herbivores (competition) and higher trophic levels (predation/mutualisms). I carried out experiments on the effects of host-plant variation on competition among co-occurring aphids, on the effects of plant variation between aphids and mutualistic interactions with honeydew-collecting ants, and on the mechanisms of sequestration of toxic cardenolides as defense of the aphid against predators. Key papers: Züst & Agrawal (2016) Functional Ecology, Züst & Agrawal (2017) Ecology |
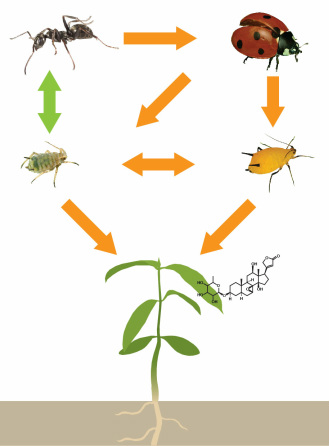
Simplified aphid foodweb for Asclepias syriaca. Orange arrows represent antagonistic interactions (predation/competition), while the green arrow represents a mutualism.
|
|
Top-down selection of plant genotypes
As part of my PhD studies at the University of Zürich under the supervision of Lindsay Turnbull, I experimentally tested the impact of herbivory by different aphid species on the genotypic composition of populations of Arabidopsis thaliana. Over five plant generations, I repeatedly exposed replicate plant populations to herbivory by the turnip aphid (Lipaphis erysimi), the cabbage aphid (Brevicoryne brassicae), the green peach aphid (Myzus persicae), or no aphids as a control. Selection by aphids mainly acted on the chemotype of the plant, i.e., the major type of defensive chemical produced by the plant. Lipaphis erysimi selected for genotypes mainly producing glucosinolates with a 3-C side chain, whereas Brevicoryne brassicae selected for glucosinolates with a 4-C side chain. Based on these results, we looked at published data on the chemotypes of natural Arabidopsis thaliana population, and found a gradient from 4-C in southwest Europe to 3-C in northeast Europe. Using long-term aphid population data provided by the EXAMINE network, we then found that this gradient in chemotypes corresponds to a gradient in the relative abundance of Lipaphis erysimi and Brevicoryne brassicae. Thus it is likely that the mechanism we found under controlled laboratory conditions is also acting at a much larger scale to shape plant populations. Key paper: Züst et al. (2012) Science |
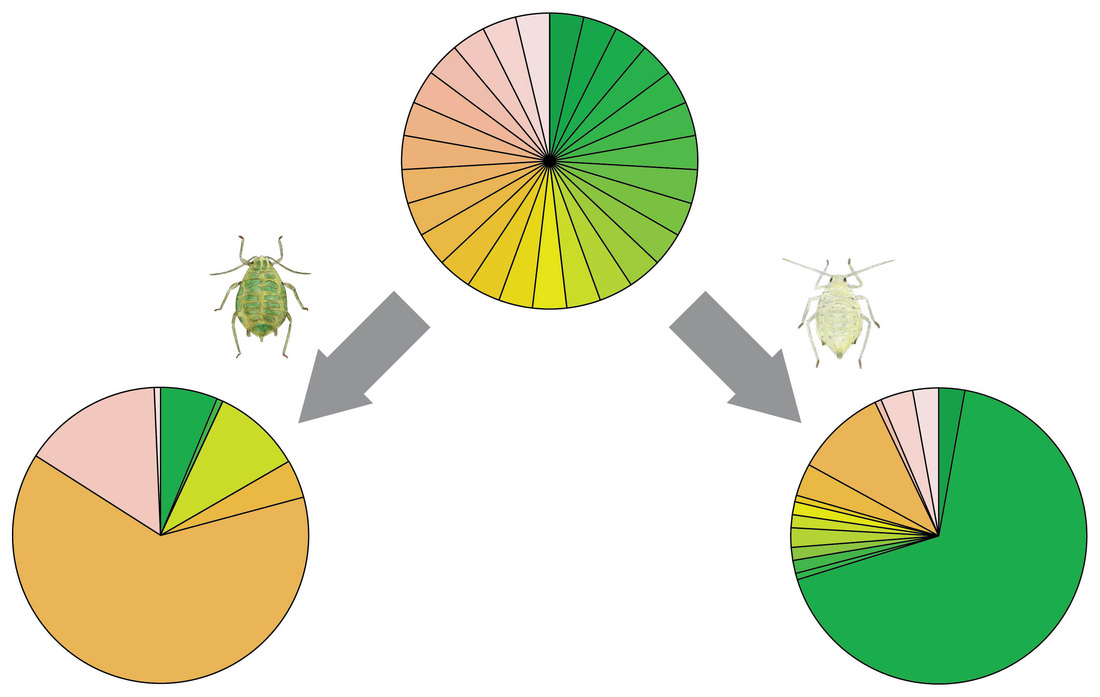
Genotype selection in response to aphid selection. Starting with 27 genotypes at equal frequencies, the aphid Lipaphis erysimi (left) selects for different genotypes than Brevicoryne brassicae (right).
|
|
Plant growth - defense trade-offs
Underlying the natural variation in plant defense is the assumption that defensive traits are costly for a plant to produce. Even though the existence of such costs has important consequences for the evolutionary interactions of plants and their enemies, their existence has frequently been challenged. While costs can manifest themselves in a variety of forms, I focus on trade-offs between plant growth and defense. I use sophisticated statistical methods to accurately describe plant growth based on repeated size measurements. This allows me to estimate physiological growth traits of plants that are often less biased than conventional size or growth measures, and thus are more likely to show true trade-offs with plant defense. Check out my paper in Annual Review of Plant Biology! |
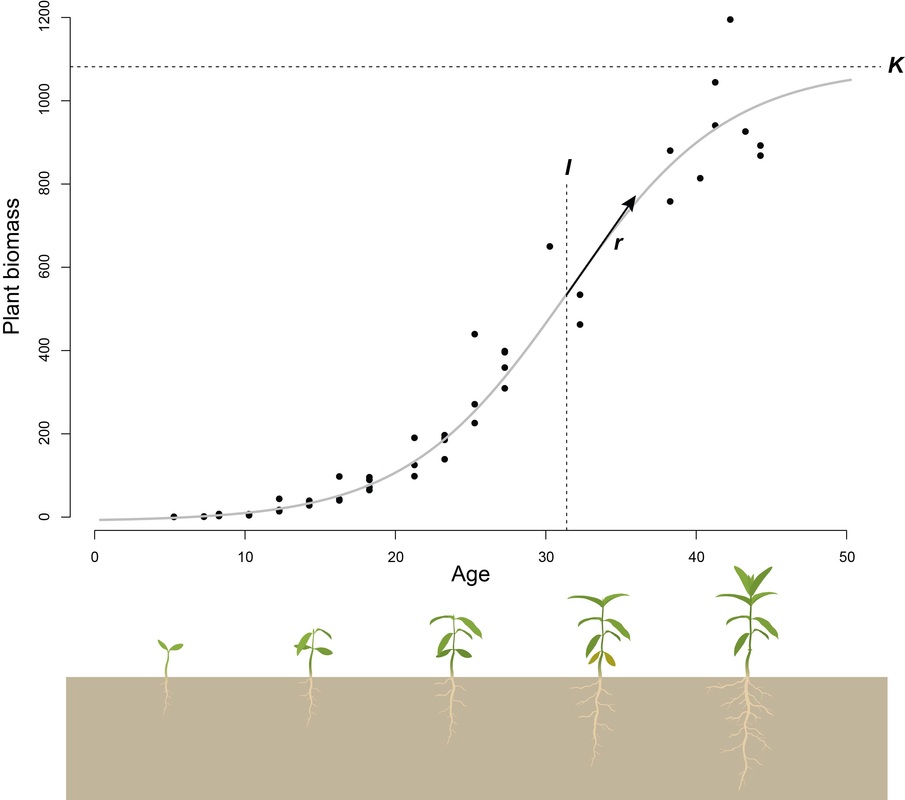
Logistic growth of plant biomass. Repeated size measurements allow the estimation of growth parameters such as asymptotic size (K) and point of inflection (I) at which the absolute growth rate (r) is maximized.
|